補体系概説
はじめに 補体はレクチン、Toll-like receptor (TLR) などと共に自然免疫系を構成する。自然免疫には特
有の識別機構が備わっており、補体も独自の異物認識機構を持つ。補体はC3bが沈着した細胞膜を後続因子で
傷害する(perforin に似た穴をあける)が、ヒト補体がヒト細胞を傷害することは無い。これはヒト細胞が
ヒト補体を選択阻害する分子(インヒビター)を発現して後続活性化を阻害するためである。ヒト細胞の代表
的補体インヒビターはCD55 (DAF), CD46 (MCP)であり、これらを表現する細胞は自己補体から守られるため、
CD46, CD55は補体の“self marker”と云える。当研究室は長くCD46研究をリードしてきた。
補体研究史は100年に亘り、そのプロトタイプは昆虫からヒトにまで保存される。補体の異物識別機構、活
性化経路に関与する因子群、自己防御に関与する制御系補体蛋白質、補体の細胞傷害活性の分子機構、補体レ
セプターとその機能、欠損症と疾患の6項目を略説する。
補体の活性化起点を提供するのはC1q またはMannose-binding lactin (MBL) である。ともに認識 ドメインとcollagen-like bundle をもつ相同分子である。C1q は抗体のヒンジ部分のアミノ酸配列を認識し、 MBLは糖鎖を認識する。糖鎖は微生物に多いmannose, GlcNAc などのクラスターであり、抗体も原則的に非自己 細胞に付くため、補体の活性化起点は異物表面に選択される。即ち抗体, MBLが付く細胞が補体にとっての異物 となる。C1q, MBLともserine protease の亜成分と共役的に働く。このprotease はC1qの場合C1r, C1s, MBLの 場合MASPと呼ばれる。C1q, MBLが異物面に付着するとC1s, MASP がそれぞれ活性化し、次項の補体protease cascade に連なる。補体系は重複性が強いのでgene duplication が起きた時代の反映と云える。
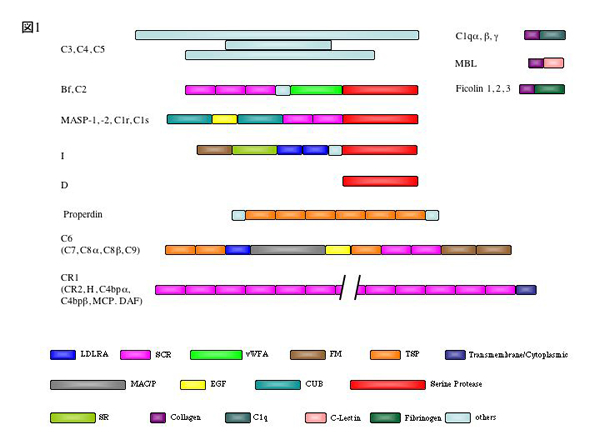
補体初期成分は血漿酵素系である。補体活性化とは異物膜上で各補体成分が所定の順番で連鎖的に 活性化して分子集合を形成しつつ種々の生物活性を発揮するプロセスである。C1s, MASPともにC2, C4 を限定分 解し、C4bC2a 複合protease を形成する。C1s, MASPは2種の補体成分を天然基質とする。C4はthioester を持ち、 活性化C4bは膜の水酸基、アミノ基などにester 結合して固定する。C4bはC2aと結合して基質C3をC3a, C3bに限 定分解する。このC4bC2a複合体(古典的C3 convertaseという)においてC2aはprotease として機能する。C3aは 血管透過性を亢進し、後述のC5aとともにanaphylatoxin と呼ばれる。C2b, C4a にも生理活性があるが省略する。 C3bには補体経路で2つの機能がある。まず、C3b自身がC4bと同様にester結合で膜にアンカーし、新たな複合C3 convertase, C3bBb を形成する。C3bBbは代替経路のC3 convertaseと呼ばれ、C3活性化を強く増幅する。次に C3bはC4bまたはC3bに共有結合し、C4bC3bC2aまたはC3bC3bBbという分子集合を形成しうる。これは基質特異性を C3からC5に変え、故にC5 convertase と呼ばれる。C5 convertase はC5をC5aとC5bに分解し、C5bは後続補体成分 C6-C9と非酵素的に複合体を形成、膜侵襲複合体(membrane attack complex, MAC)として細胞膜に穴をあける。 MACは疎水複合体であるため膜に侵入すると説明される。C5aはanaphylatoxin として血管透過性と好中球に走化 性を与える。
特記事項は補体活性化蛋白の短寿命性である。C4b, C3bの膜結合活性は1分以内に失われる。従って 90%以上の分子は膜ではなく水分子などと結合して安定化する。実際C3b, C4bはthioesterの自然乖離で徐々に失 活する。また、C3 convertase複合体も数分の寿命で乖離して失活する。これらは補体の過剰反応を防ぐためと合 目的的に説明されるが、蛋白分子の高次構造の劇的な変化を反映するものに違いなく、短時間の反応であるため に構造遷移の解析は進んでいない。
自己識別機構に関与する補体制御蛋白質補体系の細胞傷害は異物細胞に限って発揮される。自己細胞は破壊を免れる。C3b, C4bが異物膜のC1s, MASPの近くで活性化するにせよ近くの細胞(自己、非自己細胞に拘らず)に沈着することは知られており、細胞 傷害が波及した場合、reactive lysis と呼ぶ。しかし、自己細胞は決してreactive lysis を受けない。その後の 知見から、自己細胞が補体抵抗性なのは同種補体(特にC3b, C4b)を制御する分子を膜表面に発現するためと判明 した。代表的補体制御分子にはCD55 (DAF), CD46 (MCP) がある。異種細胞はこれらを持たないか、あるいはヒト 補体と互換性の無い(種の異なる)CD46, CD55 を発現するためにヒト補体によって傷害されることが判明した。 これらの知見はCD46, CD55のいづれかを表現することが同種補体から自己細胞を守る必須手段であること、言い換 えるとCD46, CD55は補体のself marker であることを意味する。
CD46, CD55 の構造的特徴はともにアミノ酸60余個から成るshort consensus repeat (SCR, or Sushi domain) を構成モチーフとすることである。補体関連のSCR蛋白はその遺伝子が1q32 に gene cluster を形成し、 Regulator of Complement Activation (RCA) gene familyと呼ばれる。RCA構成メンバーはCD46, CD55の他に後述の 補体レセプター、CR1, CR2や液性の補体制御因子factor H, C4b-binding protein (C4bp) が含まれる。Factor H, C4b-binding protein (C4bp)は共に膜上より液層のC3b, C4bに強い親和性を有し、体液中の補体の過剰活性化を防 ぐのが主な役目とされる。
補体の細胞傷害活性補体の膜侵襲(MAC)活性は後期成分の非酵素的重合による。後期成分はperforinと類似のdomainを持 ち高次構造の変化で疎水性が亢進する。MAC形成は異物膜で効果的に発揮されるが自己細胞では発揮されない。この 理由も後に明らかになった。MACの形成阻害に働く阻害因子が自己細胞上または液層にあり、これらがMACによる自己 細胞の攻撃を阻む。膜上のMAC阻害因子としてCD59, 液層の阻害因子としてvitronectin, clusterin などが知られて いる。MACは1分子づつのC5b, C6, C7, C8と約20分子のC9から成るとされる。これらの阻害因子はC9の重合を最も強く 阻害する。即ち補体の攻撃が自己細胞を回避するのはC3-step, C9-stepに天然の阻害機構があり、同種補体のみを選 択阻害するためである。
補体レセプターと細胞応答補体レセプターは広義にはC1q receptor, C3a/C5a anaphylatoxin receptor, C4a receptor, などを含 むが本項ではC3の分解産物(図1)C3b, C3bi, C3dg/C3d を認識するreceptor 群を扱う(図2)。CR1, CR2はSCRを 基本もチーフとするRCA family である。一方、CR3, CR4はbeta2-integrin である。CR5 以下の報告もあるが一般的 でないので略す。CR1,はSCR30個からなる大きい膜蛋白で樹状細胞、マクロファージに発現し、貪食促進や異物吸着に 関与する。CR1は100 コピー程度と少ないが赤血球にも発現する。赤血球CR1は免疫粘着反応の主役で、補体結合異物 をロゼット形成で肝臓などの網内系に運びクリアランスに導く主役とされる。CR2は主にB 細胞に発現し、抗原の captureと抗原レセプターへの異物分配を司る。CR2はEBVのレセプターでもある。この他、RCA family にはvirus receptor を兼ねるものが多い。
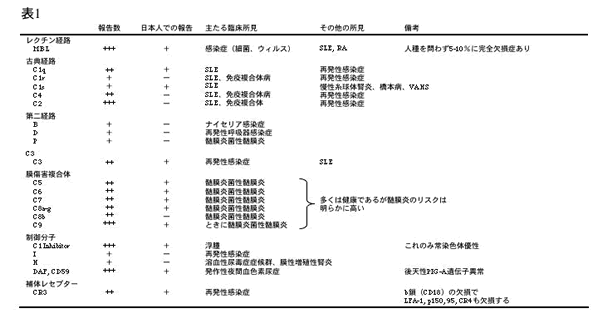
欠損症と疾患全ての補体因子(C1-C9)について欠損症が見出されたが、致死的または重篤な問題は起きない。最も多い 併発症は自己免疫疾患(特にSLE)である。これは補体系がDNA、死細胞のクリアランスに関与することを示唆する。
一方、補体制御因子、特にRCA family の欠損はHUS/TTPを誘起する例(Factor H, CD46)が報告されてい る。また、CR3/CR4 の共通欠損症はGCD という重い反復感染症を誘起する。
ブタなどにヒト補体制御因子CD46/CD55をtransgene すると、補体にとってはヒト型となり、ブタの臓器が ヒトに移植可能になる。
他に、癌、アレルギー、溶血性貧血特にPNH、多くの炎症性疾患に補体系が関与することが報告されてきた。 疾患と補体系の関連は今後の研究課題である。
- Oshiumi, H., Y. Suzuki, M. Matsumoto, and T. Seya. 2009. Regulator of Complement Activation (RCA)
gene cluster in Xenopus tropicalis. Immunogenetics 61: 371-384.
- Oshiumi, H., K. Shida, Y. Kimura, J. Katoh, S. Ohba, Y. Tamaki, T. Hattori, N. Yamada, N. Inoue, M. Matsumoto, S. Mizuno, and T. Seya. 2005. The regulator of complement activation (RCA) locus in the chicken: Identification of the chicken RCA gene cluster and functional characterization of the RCA proteins. J. Immunol. 175:1724-1734.
- Kimura, Y., N. Inoue, A. Fukui, H. Oshiumi, M. Matsumoto, M. Nonaka, S. Kuratani, T. Fujita, M. Nonaka, and T. Seya. 2004. A short consensus repeat-containing complement regulatory protein of Lamprey that participates in cleavage of lamprey Complement 3. J. Immunol. 173: 1118-28.
- Inoue, N., M. Ikawa, T. Nakanishi, M. Matsumoto, M. Nomura, T. Seya, and M. Okabe. 2003. Disruption of the mouse CD46 caused an accelerated spontaneous acrosome reaction in sperm. Molec. Cell. Biol. 23: 2614-2622.
- Fukui, A., T. Yuasa, Y. Murakami, K. Funami, N. Kishi, T. Matsuda, T. Fujita, T. Seya, and S. Nagasawa. 2002. Mapping the sites responsible for factor I-cofactor for cleavage of C3b and C4b on human C4b-binding protein by deletion mutageneis. J. Biochem (Tokyo). 132: 719-728.
- Nomura, M, M. Kurita-Taniguchi, K. Kondo, N.Inoue, M. Matsumoto, K. Yamanishi, M. Okabe, and T. Seya. 2002. Mechanism of host cell protection from complement in murine cytomegalovirus (CMV) infection: Identification of a CMV-responsive element in the CD46 promoter region. Eur. J. Immunol. 32: 2954-2964.
- Murakami H, H. Nagashima, Y. Takahagi, S. Miyagawa, T. Fujimura, K. Toyomura, R. Nakai, M. Yamada, T. Kurihara, T. Shigehisa, M. Okabe, T. Seya, R. Shirakura, and T. Kinoshita. 2002. Transgenic pigs expressing human decay?accelerating factor regulated by porcine MCP gene promoter. Mol. Reprod. Dev. 61: 302-311.
- Inoue, N., A. Fukui, M. Nomura, M. Matsumoto, K. Toyoshima, and T. Seya. 2001. A novel chicken membrane-associated complement-regulatory protein: Molecular cloning and functional characterization of a chicken membrane SCR protein. J. Immunol. 166: 424-431.
- Begum N. A., Y. Murakami, S. Mikata, M. Matsumoto, H. Hatanaka, S. Nagasawa, T. Kinoshita, and T. Seya. 2000. Molecular remodeling of human CD46 for xenotransplantation: designing a potent complement regulator without measles virus receptor activity. Immunology 100: 131-139.
- Kurita-Taniguchi, M., A. Fukui, K. Hazeki, A. Hirano, S. Tsuji, M. Watanabe, M. Matsumoto, S. Ueda, and T. Seya. 2000. Activation of human macrophages through CD46 (measles virus receptor): Production of IL-12 p40 and nitric oxide in association with recruitment of SHP-1 to CD46. J. Immunol. 165: 5143-5152.