分子進化と微生物応答

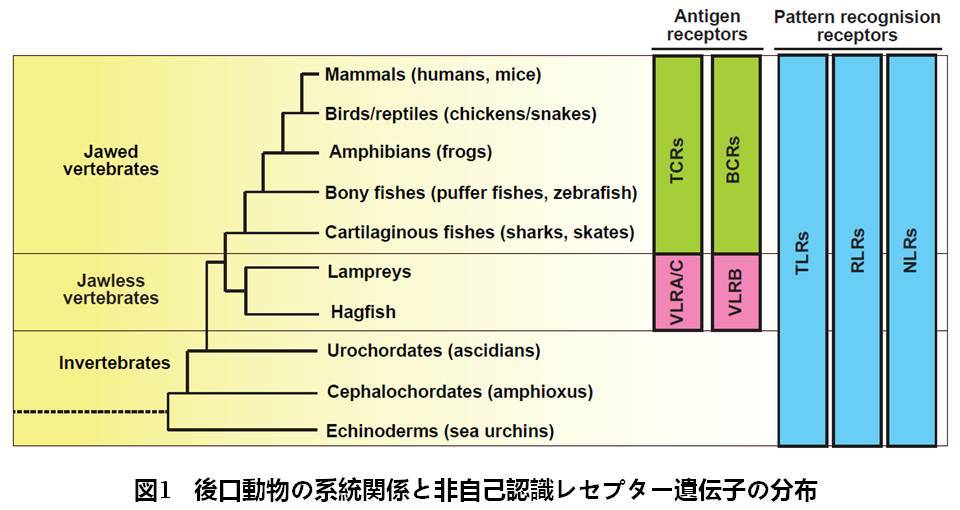
免疫系は大きく適応免疫系と自然免疫系に大別され,それぞれ異なるレセプターが病原体の認識を担います.適応免疫系ではT細胞とB細胞に発現するT細胞レセプター(TCRs)とB細胞レセプター(BCRs)が病原体の認識に与り,抗原レセプターと呼ばれます.加えて,最近の研究からVariable lymphocyte receptors (VLR)も抗原レセプターとして機能することが判明しました.一方,自然免疫系ではToll-like receptors(TLRs)やRIG-like receptors(RLRs)に代表されるパターン認識レセプター(PRRs)が微生物固有の分子構造(PAMPs)を認識します.これまでに数十種のPRRが同定されており,それぞれ特定のPAMPs を認識します.
抗原レセプターは脊椎動物のみに確認されていることから,適応免疫系は脊椎動物に固有の免疫システムです(図1).一方,PRRsは全ての多細胞生物に存在していることから,自然免疫系は多細胞生物に普遍的な生体防御機構です.そこで,私たちは様々な生物から自然免疫遺伝子を同定し,ヒトへと至る自然免疫系の進化過程を解明しています.
①TLRの分子進化と多様性
私たちは原始的な脊椎動物である両棲類(カエル)や魚類(フグ),無顎脊椎動物(ヤツメウナギ)から網羅的にTLRを同定しました.その結果,水棲環境に適応したフグやヤツメウナギにはヒトやマウスには存在しない固有のTLR(TLR14やTLR22など)が存在することが分かりました(表).不思議なことに,陸上環境にも適応したカエルは水棲動物と陸上動物のTLRをほぼ全て備えていました.暴露される病原体の種類は生活環境に依存することから,これらTLRレパトアの変化は生活環境への適応進化によって生じたことが示唆されます.

②原始TLRの機能
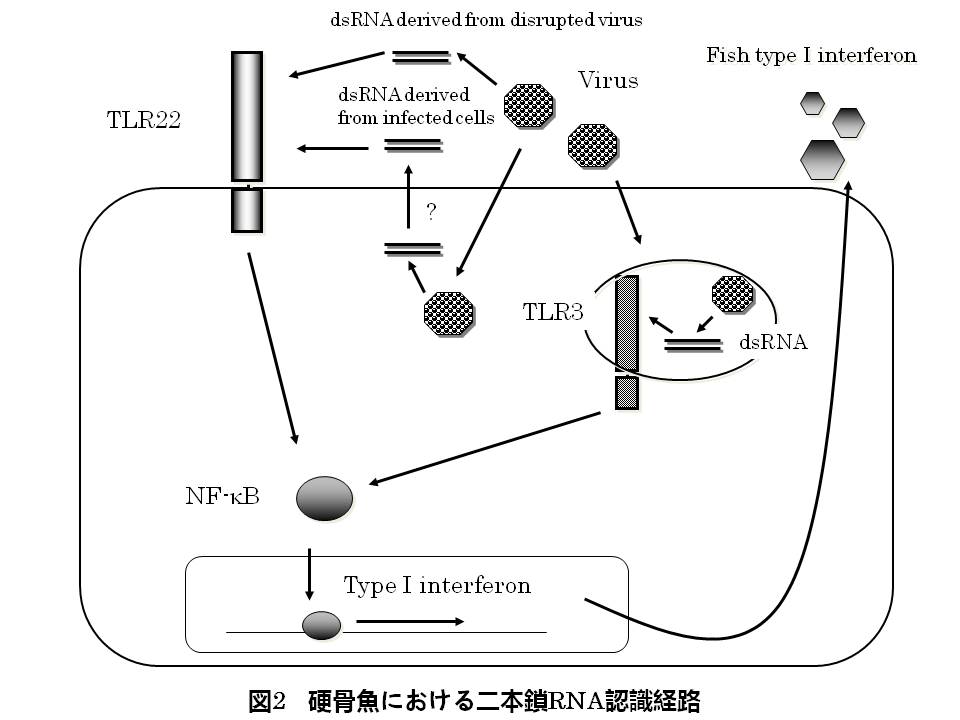
では,水棲動物に固有のTLRにはどの様な役割があるのでしょうか?一例としてTLR22の研究を紹介します.機能解析の結果,TLR22はTLR3と同様にウイルスに固有の二本鎖RNAを認識し,抗ウイルス応答を開始させることが分かりました(図2).認識するPAMPは共通ですが,魚類のTLR3とTLR22は細胞の局在や発現する組織が異なります.したがって,魚類ではTLR3とTLR22が協調して抗ウイルス応答を制御していることが示唆されます.
進化の過程で,パラサイトと宿主は共存する道(トロピズム)を選んできました.偶発的に,このトロピズムを無視する感染症が生じ,ヒトを死に導きます.その多くは鳥インフルエンザに代表される新興感染症です.自然免疫系の進化研究は生物の多様性を理解するだけではなく,これら新興感染症を制圧するために必須の基盤情報を提供します.
- Kasamatsu, J. 2013. Innate and adaptive pathogen recognition receptors in jawless vertebrates. Microbiol. Immunol. 57: 1-12 (Review).
- Kasamatsu, J., H. Oshiumi, M. Matsumoto, Kasahara, and T. Seya. 2010. Phylogenetic and expression analysis of Lamprey Toll-like receptors. Dev. Comp. Immunol. 34: 855-865.
- Seya, T., M. Matsumoto, T. Ebihara, and H. Oshiumi. 2009. Functional evolution of the TICAM-1 (TRIF) pathway for extrinsic RNA sensing. Immunol. Rev. 227: 44-53 (review).
- Oshiumi, H., A. Matsuo, M. Matsumoto, and T. Seya. 2008. Pan-vertebrate Toll-like receptors during evolution. Curr. Genomics 9: 488-493 (Review).
- Matsuo, A., H. Oshiumi, T. Tsujita, H. Mitani, H. Kasai, M. Yoshimizu, M. Matsumoto, and T. Seya. 2008. Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses. J. Immunol. 181: 3474-3485.
- Higuchi, M., A. Matsuo, M. Shingai, K. Funami, A. Ishii, H. Oshiumi, M. Matsumoto, and T. Seya. 2008. Combinational recognition of bacterial lipoproteins and peptidoglycan by chicken Toll-like receptor 2 subfamily. Dev. Comp. Immunol. 32: 147-155.
- Ishii, A., M. Kawasaki, M. Matsumoto, S. Tochinai, and T. Seya. 2007. Phylogenetic and expression analysis of amphibian Xenopus Toll-like receptors. Immunogenetics 59: 281-293.
- Ishii, A., H. Sawa, T. Tsujita, K. Shida, A. Matsuo, M. Matsumoto, and T. Seya. 2007. Lamprey Toll-like receptors with properties distinct from those of variable lymphocyte receptors. J. Immunol. 178: 397-406.
- Tsujita, T., A. Ishii, H. Tsukada, M. Matsumoto, F-S. Che, and T. Seya. 2006. Fish soluble Toll-like receptor (TLR)5 amplifies human TLR5 response via physical binding to flagellin. Vaccine 24: 2193-2199.
- Tsujita, T., H. Tsukada, M. Nakao, H. Oshiumi, M. Matsumoto, and T. Seya. 2004. Sensing Bacterial Flagellin by Membrane and Soluble Orthologs of Toll-like Receptor 5 in Rainbow Trout (Onchorhynchus mikiss). J. Biol. Chem. 279: 487588-487597.
- Oshiumi, H., T. Tsujita, K. Shida, M. Matsumoto, K. Ikeo, and T. Seya. 2003. Prediction of the prototype of the human Toll-like receptor gene family from the Pufferfish Fugu. rubripes genome. Immunogenetics 54: 791-800.
- 笠松 純,瀬谷 司,笠原 正典.「病原体認識戦略の進化」『臨床免疫・アレルギー科』,科学評論社,56(1),pp113-121, 2010.
- 瀬谷 司,松尾 綾,押海裕之.ヒトが失った微生物パターン認識機構—フグTLR解析から—,感染・炎症・免疫,医薬の門社,39 (1): 56-58, 2009.
- 石井 秋宏,瀬谷 司.「免疫応答と免疫病態の統合的分子理解に向けて」(谷口維紹・山本一彦 編) 自然免疫の分子進化,南山堂,pp23-30,2007.